![]()
Plan
Introduction Organisation générale La coquille. Influence de la torsion sur l'organisation générale. Appareil digestif Appareil circulatoire Appareil excréteur Système nerveux Organes sensoriels Appareil reproducteur Ponte et développement Systématique Sous-classe des Prosobranches
- Ordre Basommatophores
- Ordre Stylommatophores
Sous-classe des Opisthobranches
- Ordre Cephalaspidea
- Ordre Runcinacea et Acochlidiacea
- Ordre Sacoglossa
- Ordre Aplysiacea
- Ordre Pleurobranchacea (Notaspidea)
- Ordre Thecosomata
- Ordre Gymnosomata
- Ordre Nudibranchiata
- Sous ordre Doridacea
- Sous ordre Dendronotacea
- Sous ordre Arminacea
- Sous ordre Aeolidiacea
Les Gastéropodes sont des Mollusques devenus asymétriques du fait de la torsion subie par le viscéropallium, mais qui, en de nombreuses circonstances, se trouvent être secondairement symétriques. Extérieurement, c'est l'enroulement en spirale de la masse viscérale enclose dans une coquille typiquement hélicoïdale qui, très souvent, rend cette asymétrie apparente au premier coup d'œil. Toutefois, dans de nombreux cas, la coquille fait défaut à l'état adulte et les Animaux devenus limaciformes ne laissent plus discerner leur asymétrie que par la situation des émonctoires et de l'appareil copulateur au côté droit du corps. L'asymétrie des patelles et autres formes apparentées, dont la coquille est sensiblement conique, est encore moins évidente, mais elle n'en est pas moins réelle.
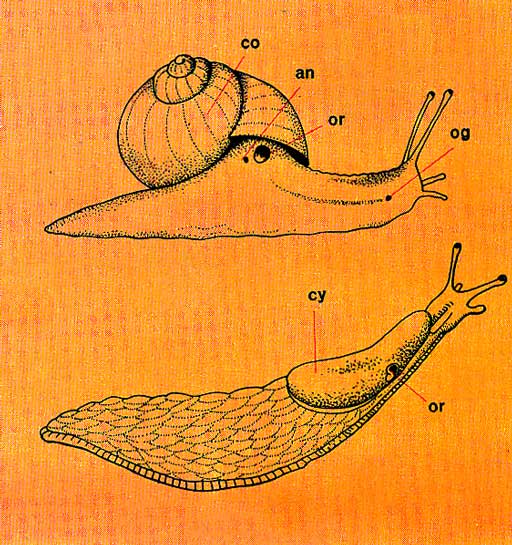
Morphologie externe. Tête, pied, viscéro-pallium, toujours présents ou presque, montrent de très grandes variations morphologiques.
La tête, souvent bien distincte du reste du corps, allongée ou non en mufle, porte la bouche et dorsalement une paire d'yeux, une ou deux paires de tentacules, ou des rhinophores. A ces organes sensoriels s'en ajoutent d'autres dont la fonction est mal connue.
Le pied demeure en général aplati en une sole pédieuse à fonction essentiellement locomotrice. C'est un organe riche en muscles, relié à la columelle par le puissant muscle columellaire qui s'y insère solidement et dont la contraction provoque la rentrée de l'Animal dans la coquille. Chez plusieurs espèces pélagiques, le pied se modifie en une large carène ou en nageoires puissantes. Ses portions latérales deviennent des parapodies natatoires lorsqu'elles s'étalent en lobes latéraux.
Le manteau existe très généralement sous la forme d'un vaste repli qui recouvre soit une cavité palléale, soit une cavité pulmonaire, mais dans plusieurs familles il disparaît pour faire place à un notum lisse ou hérissé de papilles respiratoires.
La masse viscérale est l'un des principaux éléments de la morphologie du corps. Elle s'enroule en "tortillo" ou se recourbe seulement. Elle ne fait même plus saillie dans les espèces limaciformes.
La coquille est unique. Toutefois, Kawaguti et Baba ont eu la surprise de découvrir (1959) un authentique Gastéropode à coquille bivalve, Tamanovalva limax, devenu depuis Berthelinia limax. A la suite de cette découverte, il est apparu que des valves de Mollusques actuels connues de longue date devaient être rapportées à des formes voisines de Gastéropodes.
Très tôt au cours de l'embryogénie la coquille apparaît comme une mince pellicule élaborée par le bord d'une dépression circulaire de la portion dorsale des embryons. C'est la glande coquillière. Par la suite, la coquille grandit en épousant les contours de la masse viscérale enroulée en tortillon.
L'existence de la coquille est générale chez les embryons. Toutefois, chez beaucoup d'espèces, elle disparaît lors de la métamorphose sans être compensée par une formation définitive et le corps reste nu. Dans la majorité des cas la croissance de la coquille se produit encore après la métamorphose, avec ou sans discontinuité nette.
La coquille des Gastéropodes se développe en quelque sorte sous la forme d'un hélicocône, c'est-à-dire d'une surface conique enroulée en spirale autour d'un axe hypothétique. Le pas de l'hélice est très variable et l'hélicocône peut même s'enrouler dans un plan, comme le montrent les coquilles planorboïdes. La columelle correspond à l'axe d'enroulement. Cependant, selon le degré de confluence des tours vers l'axe, elle est pleine ou creuse, et dans ce cas, ouverte à la partie inférieure par un ombilic étroit, large ou très large.
 |
 |
 |
Les tours de spire, en général délimités par un sillon plus ou moins profond, la suture, n'ont pas toujours en réalité une section circulaire. Ainsi, leur face externe devient plane dans les coquilles trochiformes. Lorsque le dernier tour, très dilaté, recouvre les autres, la suture n'est plus apparente. Dans les coquilles pupiformes la section de l'hélicocône, après avoir atteint une certaine valeur, tend au contraire à se réduire.
Le dernier tour s'ouvre par le péristome où l'on distingue le bord externe, ou labre, le bord inférieur simple, échancré, allongé en gouttière ou en tube lorsque le manteau développe un siphon respiratoire, et le bord columellaire, parfois épaissi par une callosité columellaire. Cette ouverture porte en bien des cas des plis internes, des denticules ou des lamelles qui la rétrécissent curieusement.
L'enroulement n'est pas toujours régulier ou continu.
L'hélicocône semble alors se dérouler et s'allonger comme un tube très irrégulier. Chez quelques formes terrestres, c'est peu avant le niveau de l'ouverture qu’il change brusquement de direction. La coquille des espèces patelliformes, conique, ne montre pas de tendance à l'enroulement. Plusieurs Gastéropodes du plancton élaborent un test très particulier : cône très mince, très allongé, test globuleux ou aplati des cavolinies, des Euclio.
L'ornementation des coquilles (stries, varices, cordons, tubercules, épines, lamelles) atteint un étonnant degré de complexité. Elle résulte de différences d'activité du bord du manteau qui n'a pas en toutes ses régions les mêmes propriétés.
Quelques Gastéropodes possèdent une coquille entièrement recouverte par le tissu palléal, et qui est entière ou réduite à des granulations calcaires.
Des coupes faites dans des coquilles montrent qu'elles consistent en couches superposées d'éléments cristallins agencés selon différents "types structuraux" : structures prismatiques simples ou complexes, feuilletées (nacre), entrecroisées, dans lesquelles les éléments cristallins s'ordonnent pour réaliser des ensembles d'une grande résistance.
Influence de la torsion sur l'organisation générale.
Comme on l'a vu, l'organisation des Gastéropodes est profondément affectée par la torsion du complexe palléo-viscéral et l'enroulement de la masse viscérale. A partir du Mollusque ancestral symétrique et enroulé dans un plan, on peut considérer que trois phénomènes ont eu lieu. Le premier, conséquence de l'apparition de l'axe secondaire de croissance, engendre une courbure endogastrique du complexe palléo-viscéral. En effet, l'accroissement en hauteur de cette portion du corps provoque sa déformation en U ainsi que celle du tube digestif, de telle sorte que l'anus semble se rapprocher de la bouche au-dessous de laquelle il s'ouvre. C'est la flexion ventrale.
En second lieu la portion dorsale de la masse viscérale sur laquelle s'accroît la coquille embryonnaire subit un enroulement spiralé, dit exogastrique car il s'opère dans le sens inverse de la courbure endogastrique et dans le plan sagittal, sans que la position de l'anus soit affectée, En réalité cet enroulement se poursuit soit dans le sens dextre, soit dans le sens sénestre.
Enfin intervient la torsion proprement dite, c'est-à-dire la rotation de 180° de toute la masse palléo-viscérale autour de l'axe du corps, le céphalopodium ne subissant aucune déformation. La conséquence la plus évidente de ce mouvement est le report de l'anus au côté droit puis à la partie dorsale de l'Animal. La portion du corps impliquée dans ce mouvement de rotation est celle qui se situe entre le céphalopodium, qui reste inchangé, et le complexe palléo-viscéral, l'effet de la torsion étant particulièrement net sur l'œsophage, sur le trajet du tube digestif et sur l'anse nerveuse viscérale, au moins lorsque celle-ci est suffisamment longue.
Les causes profondes de la torsion sont encore bien mal connues. Pour Garstang (1929), l'origine en serait une mutation larvaire avec pour conséquence l'asymétrie de deux muscles, l'un à gauche en rapport avec le pied, l'autre, à droite, avec la tête. La torsion débute souvent une trentaine d'heures après la fécondation. Or, à ce stade, l'organisation larvaire est encore réduite et, en fait, dans plusieurs Gastéropodes spécialisés, c'est après la torsion qu'apparaît la musculature.
Quoi qu'il en soit, une conséquence importante de ces processus est le report vers l'avant de la cavité palléale. Orientée dans le sens du déplacement, elle peut assurer ses différentes fonctions dans les conditions optimales.
Les effets de la torsion sont nets dans le groupe des Prosobranches que caractérise la streptoneurie - ou chiastoneurie -, c'est-à-dire la torsion en 8 de l'anse nerveuse viscérale. En revanche, dans les deux autres groupes de Gastéropodes, Pulmonés et Opisthobranches, l'anse nerveuse viscérale n'est pas croisée et ces formes sont dites euthyneures. Il n'en reste pas moins que la position de la cavité palléale et des orifices rénal et anal témoigne de la réalité de la torsion, parfois incomplète, ou même d'une détorsion. Il se peut en effet qu'une détorsion ait pu rétablir secondairement la situation initiale de l'anse nerveuse viscérale, mais il paraît plus probable que celle-ci n'a pas été impliquée dans la torsion par suite d'un processus de céphalisation, c'est-à-dire de regroupement des centres nerveux ganglionnaires dans la région céphalique, ce qui aurait entraîné son raccourcissement.
De la bouche, toujours antérieure, à l'anus situé en général en arrière de la tête et sur le côté droit du corps, mais parfois ouvert à son extrémité postérieure, le tube digestif comprend la cavité buccale, la cavité pharyngienne où débouchent les conduits des glandes salivaires et dont le plancher est occupé par l'appareil radulaire, l'œsophage, l'estomac et l'intestin, très long chez les formes primitives herbivores, mais beaucoup plus court chez les autres. La partie antérieure du tube digestif différencie dans plusieurs groupes une trompe évaginable, très mobile, qui se modifie chez les Pyramidellidae en un appareil de succion perfectionné. L'appareil radulaire est utilisé pour la préhension des aliments. Il est essentiellement constitué d'une radula, mince ruban porteur de rangées transversales de dents de forme extrêmement variée et dont la connaissance permet de reconnaître des genres ou même des espèces. A sa partie postérieure, ce ruban émerge de la gaine radulaire où des odontoblastes élaborent les dents, puis il passe en avant sur un odontophore, masse musculeuse souvent consolidée par des cartilages. L'odontophore, mû par de nombreux muscles, protracte la radula jusqu'au-delà de la bouche, puis la ramène à sa position initiale. La radula attaque les Végétaux ou les proies animales par un mouvement de va-et-vient. En réalité ce dernier s'accompagne de la déformation du ruban qui, d'abord creusé en gouttière, s'étale en passant sur la partie antérieure du bulbe radulaire, ce qui a pour effet de redresser les dents et de rendre leur action particulièrement efficace.
La radula n'est cependant pas toujours une "langue râpeuse". Lorsque les dents sont très longues, très fines et très nombreuses, elles agissent parfois comme une brosse ou comme un balai sur une paroi couverte de micro-organismes. Les rangées peuvent comporter de nombreuses pièces disposées avec une grande régularité, soit une dent centrale, dite rachidienne, sur l'axe du ruban, une ou plusieurs dents latérales et des dents plus externes, les marginales. Une rangée peut se réduire à la seule dent rachidienne; c'est là le type de la "radulascie" qu'utilisent les Sacoglosses pour ouvrir les parois cellulaires des Algues et aspirer le suc cellulaire. D'autres Gastéropodes possèdent de très fortes dents conformées comme des harpons et qui coexistent avec une glande à venin.
L'appareil radulaire, on le voit, se modifie considérablement avec le régime alimentaire.
L'œsophage, simple en général, différencie des glandes œsophagiennes et, chez quelques Opisthobranches, un jabot et un gésier garni de plaques ou de denticules. Par suite de la torsion il s'ouvre à la partie inférieure et postérieure de l'estomac. L’organisation de ce dernier, souvent très complexe, est en rapport étroit avec le régime alimentaire. A sa partie postérieure, le stylet cristallin est conservé par quelques groupes.
Dans l'estomac s'ouvrent les canaux de la glande digestive, masse de diverticules digestifs où semble s'effectuer la majeure partie, sinon la totalité de la digestion. Quant à l'intestin, il est long et décrit de larges anses dans les formes primitives, alors que chez les supérieures il se raccourcit beaucoup.
Au ventricule unique s'adjoignent chez les Gastéropodes les plus archaïques deux oreillettes. L'une d'elles disparaît chez ceux qui sont plus évolués. Le cœur est aidé dans sa fonction de propulsion du sang par les contractions de tout le corps, et par celles des papilles dorsales de bien des espèces dépourvues de coquille.
En liaison étroite avec l'appareil reproducteur, l'appareil rénal n'est représenté par deux reins que dans les formes primitives. De ces deux reins, le gauche, lorsqu'il n'est pas seul, est le plus important. Cette situation est évidemment l'une des conséquences de la torsion car le développement de la spire à droite du corps a provoqué, pense-t-on, la compression de l'avantdernier tour sur la partie droite de l'Animal, donc aussi une réduction de tous les organes palléaux droits.
Du point de vue de l'excrétion elle-même, on constate l'excrétion d'ammoniaque chez les espèces aquatiques et celle d'acide urique, parfois sous la forme de concrétions, chez les espèces terrestres.
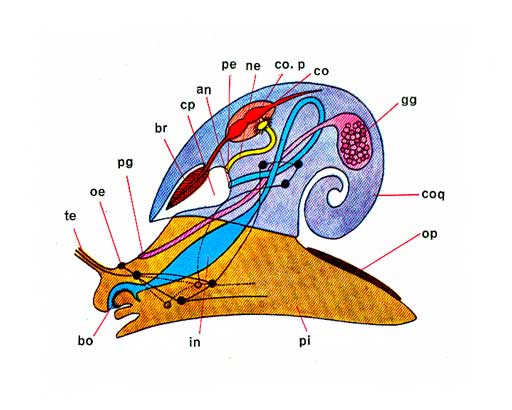
Le système nerveux des Gastéropodes montre des centres cérébraux et d'autres, secondaires, qui s'y relient par des colliers ou des anses. Mal individualisés dans les groupes inférieurs (Haliotis, Patelle...), ces centres deviennent des ganglions distincts qui, on l'a vu, tendent à se rapprocher des ganglions cérébroïdes avec lesquels certains fusionnent.
Fondamentalement, les deux ganglions cérébroïdes, dorsaux par rapport à l'œsophage, s'unissent de part et d'autre de cet organe aux deux ganglions pédieux, sousœsophagiens, reliés l'un à l'autre par la commissure pédieuse, par deux connectifs dits cérébro-pédieux. C'est là le collier périœsophagien.
L'anse viscérale, que l'on peut suivre dans la masse viscérale, se raccorde par ses deux extrémités aux ganglions cérébroïdes. Elle porte à partir du ganglion cérébroïde droit les ganglions suivants: pleural droit, pariétal droit, viscéral, pariétal gauche et viscéral gauche. Les ganglions pleuraux sont reliés aux pédieux du même côté par un connectif pleuro-pédieux.
Vers l'avant, un collier buccal porte sur son trajet deux ganglions buccaux placés sur le bulbe radulaire. Les plus archaïques des Gastéropodes montrent en outre une anse labiale passant par deux ganglions labiaux. Des nerfs issus de ces centres se distribuent à tous les organes du corps.
Les effets de la torsion se manifestent seulement sur l'anse viscérale, à condition toutefois que sa longueur soit suffisante. Les autres colliers ne sont pas affectés. Il est logique d'admettre que le ganglion pariétal droit rejeté vers la gauche au-dessus du tube digestif, est devenu le ganglion supra-intestinal et que le pariétal gauche, entraîné vers la droite, figure le ganglion sousintestinal.
Ainsi schématisée, la disposition du système nerveux peut sembler simple. En fait l'évolution de ce système pose de nombreux problèmes.
A peu d'exceptions près, tous les Gastéropodes possèdent des yeux qui peuvent être simples ou complexes, et tous sont dotés de statocystes. Par contre, des chimiorécepteurs particuliers, les osphradies, dont la fonction semble être d'apprécier les qualités de l'eau qui pénètre dans la cavité palléale, ne subsistent plus qu'exceptionnellement chez les Gastéropodes terrestres.
L'évolution de cet appareil dans toute la classe des Gastéropodes est particulièrement complexe. A la suite de la torsion il ne subsiste dans les groupes inférieurs qu'une gonade qui s'ouvre soit dans le rein droit soit dans le conduit réno-péricardique. En conséquence, les gamètes sont évacués directement en mer. Chez les autres formes, où le rein droit se réduit à son canal évacuateur, c'est dans ce dernier que débouche la gonade. C'est alors qu'apparaissent aux dépens de la partie droite du manteau des prolongements des voies génitales, sous la forme de gouttières ciliées. Un pénis se développe à droite de la tête, à l'extrémité du sillon séminal. Chez la femelle le conduit homologue devient très glandulaire. Ainsi, les voies génitales très simples des Gastéropodes primitifs subissent une spécialisation poussée par suite du développement de voies génitales palléales, puis d'organes copulateurs. D'externe, la fécondation devient interne, ce qui rend possible la reproduction des lignées devenues terrestres. Chez celles-ci du reste, comme chez les Opisthobranches qui sont tous marins, le tractus génital s'enrichit encore de glandes annexes, tandis que ses parties distales se diversifient.
Les modalités de la ponte et du développement ne sont naturellement pas les mêmes pour les formes aquatiques et pour les formes terrestres : si les premières recherchent une protection au sein des eaux, les autres utilisent les seules ressources que leur offre le sol pour mettre leur progéniture à l'abri. De même le développement des premières comporte une métamorphose plus ou moins importante alors que celui des secondes est direct.
L'œuf des Prosobranches à fécondation externe ne jouit d'aucune protection. En mer, il évolue en une larve de type trochophorien. Mais la majeure partie des Gastéropodes marins déposent leurs œufs dans des coques, des rubans ou des cordons gélatineux qu'ils fixent sur des Végétaux ou sur des roches, ce que font aussi les lymnées, Pulmonés d'eau douce. Les coques ovigères isolées ou groupées en une ponte volumineuse, telle celle de Murex brandaris, contiennent un nombre d'œufs très variable. Les œufs pondus par les Opisthobranches dans des cordons parfois très longs ou dans des rubans enroulés en spirale sont petits et abondants. On estime à plus de dix millions leur nombre dans la porte de l'ombrelle de Méditerranée (Umbraculum), à trois millions cinq cent mille ceux que dépose l'aplysie dans un cordon pelotonné de 15 ou même 20 m de long.
En mer, les larves issues d'œufs pauvres en réserves éclosent sous la forme de véligères à velum bilobé, tétralobé ou hexalobé. Elles mènent une existence méroplanctonique plus ou moins longue avant de se métamorphoser. Les Gymnosomes (Opisthobranches) ont des larves dites "polytroches" qui ressemblent à de petits tonnelets ornés de trois couronnes ciliées et se déplacent en tous sens avec une grande rapidité. Dans la curieuse larve "échinospire" de quelques Prosobranches coexistent une coquille larvaire et une coquille définitive.
Lors de la métamorphose se produit une régression ou une perte rapide du velum, mais en général les changements ne sont pas très grands. Une transformation considérable se produit par contre dans les genres Pterotrachea, Carinaria, Hétéropodes du plancton au corps effilé. En quelques heures, tandis que le velum se réduit de plus en plus, le corps, en s'allongeant énormément, change totalement d'aspect.
Le développement direct s'observe chez les Prosobranches dont les œufs contiennent beaucoup de vitellus, chez ceux qui sont incubés (Viviparus), ainsi que chez les Pulmonés. L'évolution de l'œuf de ces derniers comprend les mêmes étapes que chez les Mollusques marins, mais les organes larvaires y sont moins caractérisés. En outre, des organes. transitoires différents y apparaissent : le pied engendre à sa partie postérieure une forte vésicule pédieuse, le podocyste, ainsi qu'une volumineuse vésicule céphalique.
On admet classiquement la division des Gastéropodes en trois sous-classes, celles de Prosobranches, des Pulmonés et des Opisthobranches. Cependant, d'assez nombreux auteurs tendent à ne distinguer que deux sousclasses, celle des Streptoneures (Prosobranches) et celle des Euthyneures.
Ces Gastéropodes sont bien caractérisés par leur streptoneurie, par la présence très générale d'un opercule, élaboré par le pied, qui obture l'ouverture de la coquille lorsque l'Animal s'y retire, enfin par la séparation des sexes - à quelques exceptions près. La cavité palléale toujours orientée vers l'avant abrite deux cténidies, deux osphradies, deux glandes hypobranchiales, deux orifices rénaux chez les formes archaiques, ou un seul de ces organes.
Ecologie des Prosobranches
La répartition des Prosobranches marins est commandée par de nombreux facteurs de milieu, par le régime alimentaire et par la constitution de ces Mollusques. Ils doivent en effet pouvoir se nourrir soit d'Algues, soit de micro-organismes, soit de leurs congénères ou d'autres Animaux. Il leur faut aussi trouver une protection contre les vicissitudes du flux ou des vagues et, en fait, leurs adaptations sont très diverses. La Manche, par exemple, où les marées sont d'une grande amplitude, se prête bien à l'étude de la zonation, qui s'y révèle particulièrement nette, compte tenu des divers faciès : roches, sable, vase, herbiers. Tout en haut de la frange intertidale, dans la zone des embruns, les Littorina neritoides se groupent en petites populations dans des crevasses.
Elles sont capables de supporter une dessiccation de longue durée. Dans la zone de balancement des marées, d'autres Prosobranches doivent faire face quotidiennement à une exondation qui dure plusieurs heures vers les niveaux supérieurs. Ils se réfugient sous les pierres, sous les surplombs humides, ou dans les cuvettes, en faciès rocheux. Les cuvettes sont soumises à des variations de température très importantes. La vie n'y est possible que pour un nombre assez petit d'espèces. Les plages de sable vaseux permettent un autre moyen de protection : l'enfouissement. Beaucoup de Prosobranches y restent tapis, tout en conservant le contact avec l'extérieur par le tube du siphon et par une voie exhalante.
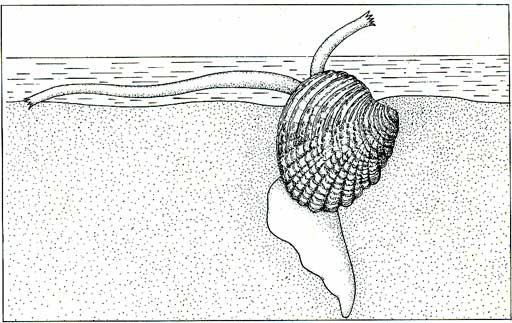
En période d'exondation, ils s'isolent dans leur coquille et n'ont plus d’activité. Dans les herbiers toujours humides que la mer laisse à découvert, les Prosobranches n'ont qu'une activité assez réduite, mais dès le retour de l'eau celle-ci reprend.
Peu d'espèces peuvent subsister sur les roches battues, mais, au nombre de ces formes, il convient de citer les patelles, au pied très puissant, qui donnent peu de prise aux vagues. L'activité des Prosobranches demeure continue sur les rivages peu affectés par les marées, ou sur le plateau continental, au-dessous de la zone des basses mers.
En eau douce, des Animaux tels que la paludine ne voient leur activité se réduire qu'en hiver. Ils s'enfoncent dans la vase et y demeurent à l'état de vie ralentie. La plupart des femelles retiennent leurs embryons dans leur oviducte jusqu'au printemps, et quand la température de l'eau dépasse une certaine limite les éclosions se produisent en masse.
Sur terre, les Pomatias elegans qui vivent normalement sous les feuilles ou à la base de différentes plantes s'enfouissent à la mauvaise saison à quelques centimètres de profondeur et ne refont surface qu'aux premiers beaux jours.
Les Prosobranches sont particulièrement intéressants par les types d'associations qu'ils peuvent constituer et par les étonnantes transformations que le parasitisme
peut entraîner chez certains d'entre eux. Les représentants de familles entières s'associent, surtout à des Mollusques et à des Échinodermes. Le parasitisme atteint les degrés les plus divers depuis les formes ectoparasites non modifiées ou peu modifiées jusqu'aux endoparasites simplifiés à l'extrême. Plusieurs Capulidae prélèvent seulement un peu de la nourriture captée par des Annélides ou des Echinodermes. D'autres, ectoparasites sur des Astéries, s'adaptent à la succion. Des Eulimidae, devenus ectoparasites sur des Astéries, ont une trompe suceuse perfectionnée.
Paedophoropus dicoelobius et Molpadicola orientalis sont endoparasites des Holothuries. La première de ces espèces a été caractérisée par ses larves véligères. Le mâle, nain, reste appliqué sur la femelle. L'appareil digestif se réduit presque à la trompe qui est longue et puissante. Parmi les Stiliferidae existent des commensaux, des ectoparasites et des endoparasites à organes très régressés (Gasterosiphon. Diacolax). La simplification atteint un degré extrême chez les Entoconchidae. Des Entocolax n'ont plus ni branchie, ni système nerveux, ni cœur, ni rein et il ne subsiste qu'un statocyste, mais l'ovaire demeure bien développé. Entoconcha mirabilis devient une sorte de Ver de 8 cm de long. L'appareil digestif des Enteroxenos disparaît en totalité. Autres parasites d'Echinodermes, les Thyonicola n'ont plus de systèmes digestif, vasculaire, excréteur ou nerveux. Le parasite se revêt de téguments appartenant à son hôte.
Le degré de simplification maximal se trouve chez Parenteroxenos dogieli, parasite de la cavité générale d'Holothuries. Cet Animal, le plus grand de tous les Gastéropodes connus, se présente comme un Ver de 130 cm de long et 4 à 5 mm de diamètre. Son corps ne contient pratiquement qu'une vaste cavité incubatrice où se voient de très nombreux cocons, un ovaire et un testicule. Les cocons donnent naissance à des larves véligères qui subissent leur métamorphose dans le tube digestif des Holothuries qui les ont avalées. La famille des Pyramidellidae et celle des Ctenosculidae renferment aussi des parasites, mais ceux-ci ne subissent pas de transformations aussi profondes.
Les Prosobranches hébergent d'assez nombreux parasites, en particulier des cercaires. Assez curieusement, l'infection massive semble provoquer une augmentation de la taille. A son degré extrême elle engendre une désagrégation de la gonade telle qu'elle aboutit à une castration parasitaire totale.
Trois grands ensembles constituent la sous-classe des Prosobranches. Ce sont les ordres des Archaeogastropoda, des Mesogastropoda et des Neogastropoda. Un quatrième ordre, celui des Neritoidea, semble devoir être séparé des Archaeogastropoda.
Ordre des Archaeogastropoda
C'est le groupe le plus archaïque, où le cœur conserve souvent deux oreillettes et où il est fréquent de trouver dans la cavité palléale deux cténidies bipectinées, deux osphradies, et une paire de glandes hypobranchiales. Les deux reins persistent en général, mais le rein droit disparaît chez les formes évoluées. La radula porte de nombreuses dents marginales. L'état archaïque se retrouve aussi dans le pied où sont développés, non pas des ganglions pédieux, mais des cordons réunis par des commissures. Les représentants de cet ordre ont un régime principalement herbivore.
Nous étudierons successivement les trois superfamilles du groupe.
Pleurotomarioïdea.
La disposition de la cavité palléale, profondément incisée, ou ouverte à sa partie supérieure, retentit sur la forme de la coquille qui est échancrée (Pleurotomaria, Scissurella), pourvue d'un orifice (Fissurella), ou de plusieurs (Haliotis) assurant la sortie de l'eau. Les pleurotomaires à coquille nacrée sont des formes exotiques de profondeur. Haliotis appelé ormeau ou oreille de mer vit au bas de la zone des marées.
Patelloidea.
Les patelles à coquille conique ont un pied sans opercule. Leurs cténidies sont modifiées en un cercle de branchies et leur radula est plus longue que le corps.
Trochoidea.
La coquille conique, turbinée, spiralée, globuleuse ou auriforme, épaisse ou mince, nacrée, est lisse ou garnie de tubercules. Trochus niloticus des mers chaudes a été très utilisé pour la fabrication des boutons de nacre. Sur nos côtes abondent les gibbules (Gibbula). Les Angaria, ou Delphinula, exotiques, ont une coquille ornée d'appendices forts, recourbés ou ramifiés, mais leur opercule est corné. Les Turbo à coquille épaisse portent un opercule épais et calcifié.
Ordre des Neritoidea
Les Nérites sont des formes marines à une seule cténidie bipectinée et au cœur traversé par le rectum. Plusieurs espèces se trouvent en eau saumâtre, en eau douce (Theodoxia fluviatilis) et même sur terre (Helicina).
Ordre des Mesogastropoda
C'est le groupe le plus important, nous étudierons successivement ses diverses super-familles.
Cyclophoroidea.
Ces Animaux terrestres ou d'eau douce à coquille souvent irrégulière ou à ornementation parfois curieuse sont presque tous exotiques.
Viviparoidea.
Les paludines de nos eaux douces (Viviparus viviparus) ne libèrent leurs embryons que lorsqu'ils sont aptes à vivre par eux-mêmes. Le tentacule droit du mâle est modifié en pénis. Les Pilidae des eaux douces tropicales (Ampullarius, Pomacea, Lanistes) sont amphibies. La partie gauche de leur manteau prend la valeur d'un vaste sac pulmonaire, mais une cténidie subsiste à droite de la cavité palléale.
Littorinoidea.
Littorina littorea, le "bigorneau", ou "vignot", abonde sur les rochers et dans les Algues. Littorina saxatilis incube ses œufs en toutes saisons. Les cyclostomes (Pomatias elegans) sont bien adaptés à la vie terrestre.
Architectonicoidea.
Les Architectonica (Solarium) de Java et de Ceylan ont un ombilic fort large laissant voir la portion axiale des tours de spire.
Cerithioidea.
Les turritelles ont une coquille très haute à nombreux tours de spire, à sculpture spirale et ouverture assez petite. Les cérithes montrent le même type d'enroulement, mais les tours portent en général des varices, des tubercules, des costulations ou des épines. Les vermets sont fixés aux roches par leur coquille à tours disjoints. Ils capturent leur nourriture par de longs filets muqueux.
Epitonioidea.
Outre les Epitonium (Scala) à coquille turriculée ce groupe contient les ianthines qui dérivent en mer grâce à un flotteur de bulles d'air confectionné par leur pied.
Aglossa.
C'est dans ce groupe que sont placées provisoirement les familles des Paedophoropodidae, des Stiliferidae, Entoconchidae, Pyramidellidae, qui contiennent les parasites déjà signalés.
Stromboidea.
Cette super-famille contient des formes ornementales telles que les Pterocera à labre étiré en longues gouttières, les Aporrhais pespelecani, les Rostellaria à labre festonné prolongé en très long rostre et même les Strombus à dernier tour plus ou moins étalé. L'œil d'Aporrhais est l'un des plus différenciés qui existent chez les Invertébrés.
Calyptraeoidea.
Outre les calyptrées (Cayptraea chinensis) en forme de chapeau chinois, ce groupe contient les crépidules (Crepidua fornicata) qui prolifèrent sur les bancs d'huîtres. D'abord mâles, ces animaux s'accumulent les uns sur les autres en courtes chaînes et changent de sexe probablement sous l'influence de sécrétions élaborées par l'individu inférieur qui est femelle.
Cypraeoidea.
Par leur galbe et leurs coloris, les cyprées sont de très beaux animaux. Ce sont les "porcelaines" à dernier tour recouvrant tous les autres et même la protoconque. Leur ouverture, très allongée, montre des lèvres régulièrement plissées. Leur manteau tacheté de vives couleurs recouvre la coquille. Il est hérissé de papilles. Sur nos côtes les Simnia ont le corps rose ou rouge et les Trivia ("pucelages", "grains de café"), de taille assez réduite, vivent sur les Ascidies.
Heteropoda.
Les Atlanta, les Carinaria et Pterotrachea sont adaptés à la vie pélagique. Les petites atlantes, à coquille discoïde, se maintiennent au sein des eaux par des battements de leur pied qui forme une longue lame très souple. Beaucoup plus grands, fusiformes et transparents, les Carinaria et les Pterotrachea ont une masse viscérale réduite, recouverte par une très mince coquille chez les premiers. Les branchies baignent largement en pleine eau. Les yeux, complexes, sont dirigés parallèlement au mufle. De l'arrière du corps des Pterotrachea part un appendice fin, plus long que le corps et dont on ne connaît pas la fonction.
Naticoidea.
Ces Animaux vivent dans le sable, et ont un pied très dilaté dans sa portion antérieure et jusque sur la tête et la coquille chez les natices. Ils se nourrissent de Bivalves (Donax, tellines, nucules) dont ils percent la coquille.
Tonnoidea.
Ce sont des Mollusques souvent de grande taille à dernier tour très ample. Tonna galea (Dolium galea) injecte par sa trompe très développée une salive toxique contenant de la tétramine qui paralyse les proies. Cassidaria echinophora n'est pas très rare en Méditerranée. Cassis madagascariensis, Cassis cornuta et les Cypraeacassis sont de grandes et belles formes. Les Cymatiidae comprennent des espèces à périostracum pileux et d'autres qui sont très grandes, comme les tritons (Charonia = Tritonium).
Ordre des Neogastropoda
Dans ce groupe la coquille est prolongée par un tube siphonal court, long ou très long, logeant un siphon bien différencié. La radula, étroite, comporte rarement plus de trois dents par rangée. Certaines espèces n'en possèdent même plus. Le système nerveux atteint son degré de concentration maximal. Le pénis, toujours du côté droit de la tête, est parfois énorme. Ces animaux sont carnivores et les procédés qu'ils utilisent pour capturer leurs proies sont variés.
Etudions les trois super-familles de ce groupe.
Muricoidea.
Communs en Méditerranée, Murex brandaris et Murex trunculus furent exploités par les Romains pour teindre des tissus. La « pourpre» de l'Antiquité provient d'une sécrétion de la glande hypobranchiale. D'abord jaune verdâtre, elle devient pourpre au soleil.
L'ornementation du test de plusieurs Muricidae consiste en longues épines (Murex tenuispina, Murex triremis), parfois aussi en longues lamelles très plissées, recourbées en gouttière (Murex palmarosae). Dans la Manche et l’Atlantique, Ocinebra erinacea dénommé se nourrit de moules et d’huîtres dont il perce la coquille. Thais lapillus, à coquille claire ou à bandes brunes, a les mêmes mœurs. Magilus antiquus se laisse emprisonner dans les Madrépores, mais au fur et à mesure de leur accroissement, il élabore un très long tube calcaire qui lui permet de survivre.
Buccinoïdea.
Les buccins (Buccinum undatum) sont fréquents dans la Manche, en bas de la zone des marées. Les nasses (Nassarius) se dirigent vers les cadavres dont elles perçoivent l'odeur à grande distance.
Volutoïdea.
Ce sont de grandes et belles formes à coquille lisse avec des zébrures sombres, qui vivent dans les mers chaudes. Voluta musica vit aux Antilles. Les harpes (Harpa major), lisses, luisantes, ont des ornementations d'une grande élégance.
Sous-ordre des toxiglossa
Mitres, cônes et Turridae (pleurotomes) sont groupés dans cet ensemble. Les mitres (Mitra papalis, Mitra episcopalis) ont une coquille fusiforme à fond clair et taches de couleur orangée ou brune. Les dessins qui ornent la coquille des cônes, belles formes à ouverture très allongée, sont d'une grande finesse. Conus (Darionicus) textilis en est un bon exemple. Plus rare, Conus gloria maris est aussi la gloire de quelques musées privilégiés. En contrepartie certains cônes sont très venimeux et la liste des victimes de Conus geographus, de Conus textilus est déjà longue. Les cônes possèdent en effet une glande à venin et des dents radulaires qui peuvent être protractées à bonne distance. Ils sont surtout piscivores et vermivores. Les pleurotomes ont également une glande à venin. Les Terebra ont une coquille très effilée à tours nombreux et petite ouverture (Terebra subulata de l'Indo-Pacifique).
Comme les Opisthobranches, les Pulmonés sont euthyneures et hermaphrodites, ce qui explique les similitudes de leur organisation. Mais leur originalité réside avant tout dans la présence d'une cavité pulmonaire à plafond très vascularisé qui ne semble pas avoir la même origine que la cavité palléale. En effet elle apparaît chez l'embryon comme une dépression ectodermique qui s'approfondit et demeure ouverte au niveau du pneumostome.
La coquille, plus souvent dextre que sénestre, n'atteint pas d'aussi grandes dimensions que chez les Prosobranches. Elle peut se réduire et disparaître. Un opercule n'en obture l'ouverture que chez les Amphibolidae.
On y distingue:
1 ° Les Basommatophores, aquatiques, dont les yeux se voient à la base des tentacules qui sont rétractiles, mais non invaginables.
2° Les Stylommatophores, terrestres, dont la tête porte en arrière de deux tentacules sensoriels deux autres tentacules rétractiles, invaginables, les ommatophores.
La tête plus ou moins nettement délimitée du pied laisse voir fréquemment l'orifice génital sur sa droite s'il s'agit d'une forme dextre, ou sur sa gauche chez les formes senestres.
La masse viscérale s'enroule en tortillon dans le cas où la coquille est spiralée, mais quand cette dernière a disparu, elle se loge dans le pied. Elle est revêtue d'un manteau apte à créer une cavité palléale (Ancylus, Chilina) où se développent chez les espèces aquatiques des branchies adaptatives (Siphonaria, Protancyius, Miratesta). Le recouvrement de la coquille par le manteau n'est pas exceptionnel et peut aboutir à la fusion des bords palléaux avec persistance d'un orifice palléal et lorsqu'il y a enfoncement de la masse viscérale dans le pied, la coquille s'isole dans une poche coquillière. Cette poche, très vaste, et d'où la coquille a disparu, recouvre tout le dos des Philomycus. Le champ palléal des espèces limaciformes demeure bien localisé. La cavité pulmonaire devient complexe chez les Arion. Elle se ramifie chez les Onchidiidae ou produit un poumon trachéen à fins tubules buissonnants chez les Athoracophoridae.
L'appareil digestif, plus voisin de la disposition primitive des Basommatophores, se spécialise. Un gésier musculeux précède l'estomac de constitution souvent simplifiée. Le cœur n'a qu'une oreillette placée en avant du ventricule. Le rein s'ouvre dans la cavité pulmonaire par une papille ou, dans les formes supérieures, par un uretère.
Le système nerveux peut se caractériser dans toute la série des formes par une tendance au raccourcissement de l'anse viscérale ou même à la fusion des centres ganglionnaires. Parmi les organes sensoriels, les yeux sont ceux qui montrent le plus de particularités. Les yeux céphaliques atteignent un haut degré de perfection. Des yeux complexes existent, sur la face dorsale des Onchidiidae.
L'appareil reproducteur comporte une glande hermaphrodite, d'où part un conduit unique qui, parvenu au niveau de la glande de l'albumine, se divise en oviducte et spermiducte distincts, débouchant à l'extérieur par deux orifices, ou encore en voies mâle et femelle unies en un spermoviducte partiellement compartimenté sur sa longueur par deux bourrelets opposés. La partie distale des voies mâles et femelles montre des différenciations multiples : diverticules, glandes, organes copulateurs parfois très complexes.
Ordre des Basommatophores
La coquille spiralée, en nacelle ou en capuchon est toujours présente. Les orifices génitaux sont en général séparés.
Ce groupe comprend des formes exclusivement marines telles que les siphonaires à coquille patelliforme et branchies situées dans une cavité palléale, des formes d'estuaire ou de rivage (Amphibolidae, Ellobiidae), ainsi que les "Pulmonés d'eau douce", c'est-à-dire les familles des Lymnaeidae, Physidae, Planorbidae, Bulinidae, Ancylidae. Bien qu'aquatiques, ces animaux respirent en général par leur poumon. Aussi doivent-ils regagner périodiquement la surface pour renouveler leur provision d'air. Lorsqu'ils parviennent au contact de l'air, leur pneumostome s'ouvre largement, s'étale, puis se referme, emprisonnant ainsi une bulle d'air. L'abaissement de la température, en réduisant l'activité métabolique, ralentit beaucoup le rythme de remontées de ces animaux. Toutefois, des adaptations particulières permettent à quelques-uns de ces Mollusques de rester immergés en permanence. Les planorbes (Planorbarius corneus) dont le sang contient de l'hémoglobine semblent tirer un parti plus grand de leur oxygène que les Lymnées. La respiration des Ancylidae doit être exclusivement tégumentaire.
Lymnées, planorbes et bulins hébergent des stades larvaires de dangereux parasites de l'homme et des animaux domestiques. La bilharziose, due à deux formes de Trématodes, Schistosoma haematobium pour la forme vésicale d'Afrique et Schistosoma mansoni pour la forme intestinale (Afrique, Antilles, Amérique), semble nécessiter le séjour des parasites par des Bulinus et par quelques Planorbidae, notamment Biomphalaria. La douve du foie, Fasciola hepatica, a pour hôte intermédiaire principal une Lymnée.
Ordre des Stylommatophores
Les Stylommatophores tous terrestres, bien caractérisés par leurs tentacules oculaires situés en arrière des tentacules sensoriels, sont extrêmement nombreux.
Leur classification, très complexe en raison de l'absence de critères bien définis, utilise dans une large mesure, outre les caractères de la coquille, ceux que peut fournir l'appareil génital : oviducte, utérus, vagin et formations annexes d'une part, spermiducte, prostate, complexe pénial d'autre part. Un sac du dard, des glandes muqueuses dites multifides représentent "l'appareil amatorial" qui contracte des rapports avec les voies mâles et les voies femelles.
Le vagin s'ouvre dans un enfoncement de la paroi du corps, l'atrium (où débouchent le conduit mâle et la glande du dard), ou plus rarement dans le sac du stylet.
Le sac du dard n'existe pas chez tous les Pulmonés. Il contient un ou plusieurs stylets utilisés lors de la copulation par l'un des conjoints qui l'enfonce profondément dans le pied de son partenaire. Le pénis, dont la définition n'est pas tout à fait la même que chez les Basommatophores, est un tube à paroi musculeuse ouvert dans l'atrium. Sa paroi interne s'orne souvent d'organes excitateurs. Les spermatozoïdes sont expulsés isolément ou plus souvent groupés en spermatophores. L'accouplement, précédé de longs préludes, est unilatéral ou réciproque.
Deux groupes très inégaux sont à considérer :
Les Gymnophila comprennent les Soléolifères, sans coquille, à manteau couvrant aussi bien la face dorsale que les parties latérales. Dans ce groupe qui contient les espèces carnivores (Rathouisia, Atopos) se trouvent les Onchidella, hôtes de la zone des marées, dont les embryons passent par le stade de larve véligère.
Les Geophila rassemblent tous les autres Stylommatophores que l'on classe surtout d'après la disposition du rein, de l'uretère et du pore urinaire. Plusieurs familles telles que les Pupillidae, les Enidae ou Bulimulidae ont une coquille petite ou de taille moyenne. Les Partulidae, des îles du Pacifique, sont ovovivipares. Ils vivent sur les buissons ou même sur les arbres. Les Clausiliidae n'ont pas d'opercule, mais une lame mobile, le clausilium, en tient lieu. Les Strophocheilus, de taille assez grande, pondent des œufs très volumineux. Les succinées vivent dans les pièces d'eau ou sur les Végétaux qui les entourent. Les Tracheopulmonata n'ont plus en guise de coquille que des corpuscules calcaires inclus dans le tégument dorsal. Ils ont un poumon arborescent ou trachéal. Les Achatinidae, de taille parfois importante, sont bien connus. Achatina fulica cause des dommages considérables aux cultures, dans les régions chaudes où on a introduit ce Mollusque en tant qu'animal comestible. Les Arionoidea à coquille discoïde, carénée ou héliciforme, déprimée, ont des génitalia simples. Mais leur coquille se réduit parfois beaucoup. Le manteau recouvre alors la partie antérieure du corps. Arion rufus, la grosse limace commune au bord de nos pièces d'eau, et Arion ater sont très fréquents sous nos climats. La superfamille des Limacoïdea rassemble des formes à coquille spiralée ou réduite à une plaque calcaire (Limacidés) ou encore à coquille rudimentaire ou absente. Limax maximus, Agriolimax agrestis, et Milax gagates des régions méditerranéennes nous sont familiers. Les testacelles appartenant à une superfamille voisine sont très allongées, et portent une petite coquille sur l'extrémité postérieure de leur corps. Ces animaux voraces, capables de s'enterrer très profondément, pourchassent les lombrics. Les oléacines, également carnivores, s'attaquent à d'autres Pulmonés. Enfin, la très vaste superfamille des Helicoïdea, à coquille héliciforme, discoïde, lentiforme, conique ou ovoïde, a une très vaste dispersion. La famille la plus représentative contient nos escargots comestibles : Helix pomatia, l'escargot de Bourgogne à grosse coquille claire, et Cryptomphalus aspersa, escargot des vignes, ou "petit gris", à coquille flammulée, introduit dans toutes les parties du monde. Tous ces animaux commettent des dégâts dans les cultures potagères.
Les Opisthobranches n'ont aucun représentant adapté à la vie sur terre. Bien qu'elle soit assez nettement délimitée, cette sous-classe est très diversifiée. Les tendances évolutives y sont très marquées de sorte qu'il est bien difficile de fournir un plan d'organisation de valeur générale.
L'un des caractères les plus importants concerne la position relative du cœur qui, dans son péricarde, comprend en général en avant de la branchie un ventricule antérieur par rapport à l'oreillette ou placé sur sa gauche. L'euthyneurie est évidente, bien que certaines formes inférieures conservent un système nerveux typiquement streptoneure.
L'aspect de la tête peut être très modifié par le développement d'un fort "bouclier céphalique" et par les rhinophores qu'elle porte parfois. Le pied lui-même s'adapte de plusieurs façons à la natation, mais en général il demeure un organe de reptation développé à des degrés divers. Le manteau conserve parfois ses caractères. Toutefois il est assez fréquemment remplacé par un notum. La cavité palléale parfois développée abrite une cténidie, une glande hypobranchiale, l'anus et le néphroprocte. Elle devient virtuelle dans des groupes entiers, ce qui entraîne la disparition de la branchie et son remplacement par des branchies de néoformation, localisées en certains points du corps, ou par des papilles dorsales. Plusieurs familles ne possèdent pas de formations pouvant être interprétées sûrement comme des organes respiratoires, mais il est certain que la respiration peut être exclusivement tégumentaire.
La coquille, solide ou fragile, est spiralée, dextre ou sénestre, ou ovoïde, patelliforme, conique, réduite et incluse dans le manteau, ou absente. Elle comprend deux valves chez Berthelinia et devient aberrante dans le groupe des Thécosomes pélagiques. L'opercule est assez rarement présent.
L'appareil digestif, adapté à des régimes alimentaires variés, différencie un jabot, un gésier à plaques masticatrices dites "gésiales" et un estomac qui, bien que parfois assez spécialisé, semble n'être qu'un lieu de transit des aliments vers la glande digestive. Mais en relation avec les régimes alimentaires différents, les organes de préhension des aliments, la radula et d'autres formations annexées à la bouche ou au pharynx deviennent très spécialisés. La situation de l'anus sur le côté droit du corps, ou en arrière et dans le plan médian, pourrait résulter d'un processus de détorsion.
L'appareil reproducteur atteint un très haut degré de complexité chez bien des Opisthobranches. Le conduit mâle relié à une prostate aboutit à un pénis situé du côté droit du corps. L'oviducte, qui se complique d'une glande de l'albumine et de glandes nidamentaires, s'ouvre à l'extérieur par un orifice placé en arrière du pénis (il y a alors diaulie), mais lorsque le conduit hermaphrodite aboutit à un orifice commun (monaulie), les spermatozoïdes sont dirigés vers l'organe copulateur mâle par un sillon séminal cilié. Lorsque l'orifice de l'oviducte et le vagin sont distincts il y a triaulie.
Le système nerveux montre une évolution remarquable, car tous les intermédiaires existent entre le système nerveux à anse viscérale longue et ganglions bien distincts et celui de certains Nudibranches où les ganglions du collier œsophagien fusionnent en une masse unique avec ceux de l'anse viscérale. Les yeux, contrairement à ceux des Prosobranches, sont en général enfoncés dans le tégument et réduits. Quelques espèces en seraient dépourvues. Les "organes de Hancock" propres aux Opisthobranches doivent être gustatifs ou olfactifs. Ils se localisent aux côtés du bouclier céphalique. Il se pourrait qu'ils soient à l'origine des rhinophores, organes sensoriels développés en arrière des tentacules.
A deux exceptions près, Strubellia paradoxa et Acochlidium amboinense, qui peuvent exister en eau douce, les Opisthobranches vivent sur les tonds marins ou dans le plancton et sont surtout nombreux dans les mers chaudes. Peu d'espèces habitent les eaux saumâtres.
Ordre des Cephalaspidea
La coquille, bien constituée chez les Acteon Hydatina, Bulla, régresse plus ou moins dans plusieurs familles. Sur la tête se différencie un bouclier céphalique bien délimité, saillant, où les yeux ne sont pas toujours très visibles. La branchie est ici un repli du feuillet interne du manteau qui se plisse parfois énormément, surtout chez Acera (Akera). Sa situation par rapport au cœur, au rein et à l'anus semble représenter divers stades de la détorsion et du changement du cœur. Dans le gésier, aux plaques gésiales peuvent s'ajouter des denticules.
L'organisation de l'Acteon est très proche de celle des Prosobranches. Les Bulla, à coquille ovoïde, se trouvent dans presque toutes les mers. Leur pied n'a plus d'opercule. Gastropteron est un bon nageur grâce à son pied modifié en larges parapodies. On trouve sur nos côtes des Scaphander et des Philine.
Ordres des Runcinacea et des Acochlidiacea
Ces deux ordres comprennent de petites formes. Philinoglossa n'a qu'un millimètre de long.
Ordre des Sacoglossa
Outre Berthelinia à coquille bivalve, on classe chez les Sacoglosses Lobiger, aux parapodies allongées en deux paires de longues lames natatoires, Oxynoe et des formes telles que Hermaea, Stiliger, Alderia , à papilles dorsales foliacées ou fusiformes qui manquent chez Elysia et Limapontia. La radula des Sacoglosses se réduit à la série des dents rachidiennes, mais les dents usées, au lieu d'être éliminées comme ailleurs, s'accumulent dans un caecum placé en avant de l'odontophore.
Ordre des Aplysiacea
D'assez belle taille, les aplysies, qui présentent une tête allongée suivie d'une portion viscérale plus dilatée, sont bien caractérisées par leurs larges parapodies qui, en ondulant, leur permettent de nager. Inquiétées, elles rejettent une sécrétion violacée bien connue. Leur accouplement que l'on peut observer au printemps, du fait de la réciprocité des fonctions, donne lieu à la constitution de chaînes d'une dizaine d'individus dont seuls les extrêmes n'assurent qu'une fonction. Les Dolabella et les Dolabriferidae vivent dans les mers chaudes. Les Notarchus, de Méditerranée, effectuent de légers sauts par expulsion brusque de l'eau de leur cavité palléale.
Ordre des Pleurobranchacea (Notaspidea)
Trois types d'animaux constituent cet ordre : les Umbraculidae et les Tylodinidae, à coquille externe, ainsi que les Pleurobranchidae, dont la coquille est externe, interne ou absente. L'ombrelle de la Méditerranée (Umbraculum mediterraneum) se reconnaît à son énorme pied verruqueux arrondi et à sa coquille surbaissée d'où émerge une grande branchie. Bien que cette espèce ponde à chaque saison plus de dix millions d'œufs, il semble qu'elle soit en voie de disparition. Les tylodines, de Méditerranée, ont un corps jaune soufré et une coquille patelliforme. Le "pleurobranche" Berthella plumula n'est pas rare dans la Manche.
On a longtemps groupé tous les Mollusques planctoniques possédant des nageoires sous la dénomination de "Ptéropodes". Or ces Animaux se répartissent en deux ordres distincts : les Thécosomes, dérivés des Cephalaspidea, et les Gymnosomes, certainement apparentés aux Aplysiacés.
Ordre des Thecosomata
Chez les Euthécosomes, le pied, ventral, se situe en arrière de la nageoire qui consiste en deux lobes latéraux. La coquille est spiralée, ou très modifiée. Les Euthécosomes ont leur pied déplacé vers l'avant et différencié en une trompe plus ou moins longue. La nageoire insérée ventralement par rapport au pied s'étale en une pièce unique. Les petits Spiratella ont conservé une coquille spiralée, mais le test des Diacria et Cavolinia semble formé par l'accollement de deux valves lisses, striées, ou costulées. Les Creseis à coquille subulée abondent dans le plancton.
Parmi les Pseudothécosomes, Cymbulia peroni est l'une des formes dotées d'une pseudoconque, qui est d'ailleurs très élégante. Corolla et Gleba lui sont apparentés. Desmopterus ne possède plus aucun test. Les Pseudothécosomes, au corps presque transparent, sont de très belles créatures adaptées à la vie pélagique.
Ordre des Gymnosomata
L'adaptation encore plus poussée à la vie pélagique se retrouve chez les Gymnosomes dont le corps fusiforme et plus robuste reste de dimensions plus petites. La cavité palléale a disparu, mais des plissements tégumentaires doivent correspondre à des branchies de néoformation. Les organes de préhension des proies sont particulièrement remarquables. Le vestibule buccal s'évagine en une trompe simple ou complexe pouvant porter une mâchoire et des "sacs à crochets", sortes de dépressions d'où émergent des dizaines de crochets longs et fins. Lorsque la trompe est relativement peu développée, elle porte à sa base des "cônes buccaux". La trompe des Cliopsidae atteint deux ou trois fois la longueur du corps. Celle de Pneumoderma atlanticum porte une gigantesque paire de "bras à ventouses" rappelant ceux des Céphalopodes. Seul le bras médian est développé chez Pneumodermopsis canephora et porte une unique ventouse qui est énorme. A l'état de repos toutes ces productions sont ramenées à l'intérieur de la région céphalique. Les cliones (Clione limacina) se groupent fréquemment en essaims. Leur corps mesure de 30 à 40 mm de long. Communs en mer du Nord et dans l'Arctique, ils sont consommés par les baleines. Dans l'Antarctique, cette espèce est remplacée par Clione antarctica.
Ordre des Nudibranchiata
En dépit de nombreuses différences d'organisation, les Nudibranches constituent un ensemble de Gastéropodes que l'on peut répartir en quatre catégories relativement homogènes dont Odhner a fait quatre ordres : Doridacea, Dendronotacea, Arminacea, Aeolidiacea. Tous marins et dépourvus de coquille à l'état adulte, ces animaux ont l'aspect de limaces. Leur pied est presque toujours aplati en sole et leur notum est nu ou papilleux. La tête porte des tentacules et des rhinophores. Les organes respiratoires sont représentés par de vraies branchies ou par des lamelles latérales, des papilles ou des productions arborescentes. La glande digestive est massive ou ramifiée. Ses diverticules peuvent s'introduire jusque dans les rhinophores et les papilles dorsales. L'anus est médio-dorsal ou sur le côté droit.
Nous considèrerons les quatre groupes de Nudibranches comme des sous-ordres :
Les Doridacea ont un corps plutôt aplati. Il porte des branchies médio-dorsales disposées en rosette autour de l'anus. Doris verrucosa, Chromodoris elegans et Archidoris tuberculata vivent sur nos côtes.
Les Dendronotacea comprennent Tethys leporina qui porte un large velum antérieur, et les Phylliroe pélagiques à corps comprimé latéralement et scintillant à l'obscurité.
Les Arminacea montrent des lamelles branchiales sous le notum qui est souvent sculpté longitudinalement.
Les Aeolidiacea portent des papilles dorsales disposées en séries ou en groupes. Le corps mince est extrêmement allongé chez plusieurs espèces. Aeolidia papillosa, de couleur chamois, est commun au printemps sur toutes nos côtes.
Pour me contacter, me faire part de vos idées, me poser vos questions, me laisser vos remarques...
cliquez ici :

![]()
Retour vers le sommaire
![]()