![]()
Sur la Piste de la Molécule du Vivant
Retour il y a 6,0 milliards d'années. Réunion au somment dans l'eau Des molécules de plus en plus complexes L'atmosphère, les sources chaudes et l'espace... Des molécules à la vieLa Terre vient de naître et les océans viennent de se former...
Le temps passe encore et après des millions et des millions d'années, les premières matières organiques se forment dans le lit des océans mais faute d'oxygène et donc de toute combustion, elles s'accumulent en quantités gigantesques, réagissent les unes sur les autres, produisent des corps toujours plus complexes, préparent les matériaux de la vie.
C'est l'importance de quelques atomes (C,H,O,N, P, S ou Carbone, Hydrogène, Oxygène, Azote, Phosphore et Soufre) dans la soupe primitive qui est à la base de toutes les molécules organiques à venir. L'argile et les nombreux sels minéraux ou autres substances dissoutes provenant des continents et de l'atmosphère s'accumulent dans les eaux encore claires des océans. Bientôt toutes ces matières minérales saturent les eaux pour former la soupe primitive. Sous l'action de diverses formes d'énergies les matières minérales vont former les premières molécules organiques :
- la chaleur : issue des laves, sources hydrothermales et des rayonnements solaires, fourni une partie de l'énergie nécessaire à la transformation de matière minérale en matière organique, elle favorise également l'agitation moléculaire qui est essentielle aux rencontres et donc aux réactions.
- L'électricité : La foudre, provenant des mouvements atmosphériques, a participé à la création de nouvelles molécules.
- Le rayonnement UV : Ces rayonnements solaires, très nocifs, ont permit de nombreuses synthèses de molécules organiques en permettant la libération d'électrons de certaines molécules et leur utilisation dans les réactions biologiques..
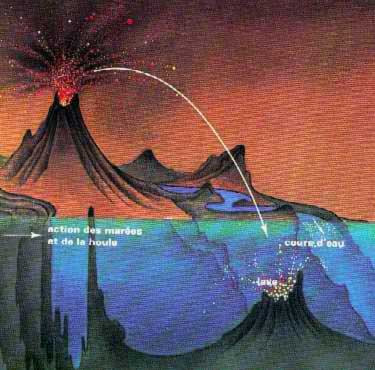
Toutes les molécules ainsi formées ont été réunies dans l'océan primitif sous l'effet des pluies. Une fois dans l'eau, les molécules se sont trouvées protégées du rayonnement UV.
Ces rayons ont aussi une action destructrice envers des molécules peu stables ou longuement exposées. Cette protection n'a lieu qu'à une certaine profondeur. Cela a permit aux molécules de bénéficier d'un "gradient" d'énergie pour pouvoir s'associer (par hydrolyse ou grâce à la présence de catalyseurs comme la chaux). Des molécules organiques simples sont ainsi apparues à cette époque :
- Certains acides aminés (qui sont à la base des protéines) déjà présents dans les comètes qui ont bombardé la Terre pendant des millions d'années,
- Certains oses, comme le ribose formé à partir du formaldéhyde, (qui sont à la base des sucres ou glucides),
- Certains acides gras, (qui sont à la base des graisses ou lipides)
- D'autres molécules importantes comme les thioesters, formés à partir d'H2S, les bases puriques (comme l'adénine formée à partir d'HCN), puis certains nucléotides...
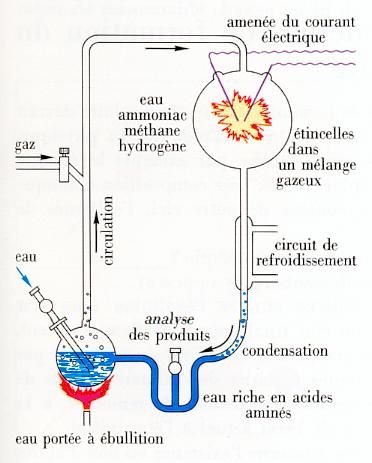
En 1953, Stanley J. Miller va faire une expérience pour tenter de comprendre comment de telles molécules ont pu se former. Il recrée les conditions atmosphériques et océaniques supposées de l'époque dans une enceinte hermétique. Il simule les éclairs avec des décharges électriques. Au bout d'une semaine d'expérience une substance rouge apparaît au fond du ballon. Elle comporte des acides aminés, ces éléments qui sont les composants de la vie. Stanley Miller provoque la stupeur mais la preuve est faite : ce lien entre l’inerte et le vivant existe bel et bien, c’est le trait d’union entre les composants de l’Univers et nous. On essaie de transformer cette matière extraterrestre (venant des comètes) et la matière minérale en matière vivante en laboratoire. Pour l’instant, on n’a pas réussi encore. C’est très difficile car il a sans doute fallu sur la terre primitive des centaines de millions d’années pour partir des composants simples et arriver à la vie. Et ça, avec notre pauvre durée de vie de scientifique, on ne peut pas le reproduire expérimentalement, évidemment.

Reste un problème de taille : les acides aminés sont bien les briques du vivant, mais ce n’est pas encore la vie.
Des expériences plus complexes montrèrent par la suite que la quasi-totalité des acides aminés, certaines bases azotées (qui entrent dans la composition du matériel génétique) et des sucres pouvaient être créés dans des conditions semblables. Les résultats de cette expérience deviennent beaucoup moins probants si l'on utilise la composition atmosphérique telle qu'on la suppose actuellement.
La présence de nombreux minéraux métalliques dissous dans les océans (Fe, Mg, Mn, Cu, Ca, Mb, Zn, Co) a probablement aidé à catalyser les réactions de formation des molécules organiques. Et ce, en particulier dans les zones riches en argile (milieu de faible profondeur) et de pyrite (milieu profond) où ces éléments permettaient de "maintenir" temporairement les molécules en relation facilitant les réactions et l'action des catalyseurs. Rapidement les dépôts de molécules organiques, sur les fonds marins et lacustres, ont formé une couche épaisse où les interactions chimiques entre molécules ont permis l'apparition de nouvelles espèces moléculaires.
Certains peptides ont ainsi pu se former par l'assemblage de quelques acides aminés entre eux, de même que des thioesters d'acides aminés, pouvant posséder une activité catalytique propre. Parmi la diversité des molécules formées, certaines se sont associées, augmentant ainsi leur stabilité, Rapidement l'océan s'est enrichi de nouvelles molécules plus complexes.
La grande majorité des réactions permettant l'apparition de molécules organiques nécessitent la perte d'un oxygène lors de l'assemblage des 2 molécules précurseurs. Or cette libération d'oxygène se fait en milieu aqueux et nécessite en fait la libération d'une molécule d'eau. Or il est impossible qu'une réaction de ce type se fasse spontanément en milieu aqueux (comment libérer de l'eau dans un milieu qui en est déjà sursaturé !). Pour qu'elle ait lieu, il faut forcer la réaction et donc fournir de l'énergie. Cette énergie sera fournie entre autres par le rayonnement UV qui va agir sur le fer dissout de l'eau (en association ou non avec du H2S dissout) et permettre la libération d'électrons à haut potentiels en énergie. Électrons qui vont immédiatement se combiner aux protons (H+) libres dans l'eau (surtout si l'on suppose le milieu acide) pour former de l'hydrogène moléculaire (H2). Hydrogène qui va permettre de forcer l'oxydation des molécules par hydrolyse et qui donc permet à la réaction d'avoir lieu ! La présence des thioesters et de phosphore sous forme de pyrophosphates inorganiques va également favoriser les synthèses de molécules organiques. Ces deux types de molécules permettent en effet d'échanger l'eau libérée contre de l'énergie. Ainsi le principal problème de synthèse de molécules organiques en milieu aqueux est résolu.
L'accumulation des premières molécules organiques va favoriser leurs rencontres et la formation de composés plus complexe par association. C'est donc principalement dans les milieux très concentrés en ces molécules que ces synthèses ont eu lieu. Ainsi les mares, les lacs et même les flaques ont joué un rôle non négligeable dans l'apparition de la vie !
De même certaines substances minérales ont favorisé des réactions. L'argile par ses propriétés physico-chimiques a facilité la polymérisation de certaines molécules. D'autres substances ont joué le rôle de catalyseurs. N'oublions pas que ce sont des milliards de milliards de molécules qui ont pu interagir ensemble grâce à l'agitation des eaux, et ce, pendant des dizaines de millions d'années. "La soupe chaude primitive" comme disent les spécialistes est prête...
Au cours des époques, on a successivement imaginé que la vie se générait spontanément ou bien émanait d'une "force vitale" imprégnant la matière. Certes, personne n'a encore aujourd'hui réussi à donner naissance en laboratoire à des "humoncules", ces humains miniatures que les alchimistes pensaient obtenir grâce à la pierre philosophale. Mais l'expérience fondatrice de Miller a déclenché une foule de travaux en chimie pré biotique, celle qui étudie les réactions ayant pu donner naissance à la vie sur Terre, il y a environ 4 milliards d'années. Les premières manifestations vitales dûment identifiées remontent à cette époque.
Les molécules du vivant sont souvent des assemblages longs et complexes d'atomes. La démarche de la chimie pré biotique s'apparente à un jeu de Meccano : elle part d'atomes ou de molécules très simples faites de carbone, d'hydrogène, d'oxygène, d'azote, de phosphore... puis elle fabrique des molécules-briques de base, composées de quelques dizaines d'atomes, comme les acides aminés qui constituent les protéines.
Seule une molécule permettant de diriger la formation d'autres molécules complexes a pu ouvrir la voie à de nouvelles réactions et combinaisons qui peu à peu ont permit l'apparition des protéines, glucides et lipides. Cette molécule, c'est l'ARN.
Parmi les molécules formées dans la soupe primitive, apparaît l'AMP, une association entre l'adénine (une base azotée), un ribose (un sucre) et un pyrophosphate inorganique (Pi). Ce dernier a d'ailleurs dû jouer un rôle dans la formation et l'accrochage du ribose à l'adénine.
L'association adénine et ribose est appelée Adénosine et forme un nucléoside (qui est le composant de base des acides nucléiques). Une fois l'AMP formé, par addition avec 2 autres pyrophosphates, il y a formation d'ATP. Cet ATP possède les caractéristiques des thioesters (il échange de l'énergie, en libérant un pyrophosphate, contre une molécule d'eau) mais il peut aussi former de longues chaînes, permettant de créer une réserve stable de pyrophosphates, par polymérisation : ATP + ATP -> ATP-AMP + Pi ATP + ATP-AMP -> ATP-AMP-AMP + Pi etc... Cette chaîne ATP-AMP-AMP-AMP-AMP... est plus connue sous le nom de PolyA. Peu à peu d'autres bases azotées sont apparues dans la soupe primitive au hasard des réactions : cytosine, uracile, guanine, et autres... Possédant les mêmes propriétés que l'adénine, ces bases ont formé du CMP, UMP, GMP et autres... qui se sont insérés naturellement dans les PolyA, formant ainsi un ARN primitif. A ce stade l'ARN ne possède encore aucune information, ce n'est qu'une succession de nucléotides. Parmi les bases azotées formées seules la guanine, la cytosine, l'uracile et l'adénine ont été conservées. Sûrement car ces molécules pouvaient se lier entre elles : La (G) se lie à la cytosine (C), l'uracile (U) à l'adénine (A), se protégeant mutuellement des dégradations.
Cette propriété, en plus de stabiliser la molécule d'ARN, a permit à celle-ci de se replier sur elle-même : certaines séquences étaient complémentaires d'autres et se liaient entre elles.
Ces repliements en boucle permettent l'apparition de nouvelles propriétés : une meilleure résistance à la dégradation par exemple mais surtout la possibilité d'une réplication de la molécule avec la formation d'un second brin complémentaire du premier. En effet, imaginons une molécule d'ARN dont une extrémité se replie. De nouvelles bases vont pouvoir s'accrocher à cette extrémité mais, en raison de leur affinité, leur mise en place ne sera plus aléatoire mais conditionnée par la base située sur l'autre brin. Parmi les différents ARN formés certains possèdent une activité catalytique (on les appelle des ribozymes). Cette activité leur fournit entre autre une capacité d'autoréplication.
Un ARN qui peut se répliquer, peut donc s'amplifier, au détriment des autres ARN (qui utilisent les même bases azotées) : c'est le début d'une sélection de type darwinienne. Les nombreuses erreurs de réplication permettent de créer de nombreuses variantes de cet ARN dont certaines seront plus efficaces (ARN plus stable, réplication plus rapide..). Par ce phénomène, où seules les bases azotées A, C, G, U pouvaient intervenir, la disparition des ARN composés avec d'autres molécules que les 4 bases azotées était inéluctable.
D'autres hypothèses font intervenir des molécules plus petites, de constitution proche des acides nucléiques, qui par leurs propriétés vont être les précurseurs des ARN. Il y a le modèle des APN (acide peptide nucléique) où ce sont de petits peptides qui portent les bases azotées ou le modèle du p-ARN où le ribose, supposé rare, est remplacé par du pyranose.
Il y a une dizaine d'années, des chercheurs ont fait une découverte de poids qui fait incliner la balance vers l'option de l'apparition première de l'ARN : sous certaines conditions et pour certaines de leurs tâches, les molécules d'ARN sont capables de se passer des protéines-enzymes. Il est donc possible d'imaginer un monde vivant n'ayant pas recours aux protéines, uniquement basé sur l'ARN, dont la construction devient un problème central de la chimie pré biotique. Car le scénario vers la vie est déjà bien ébauché, notamment le passage à la cellule : les chercheurs ont remarqué qu'en milieu aqueux, les grosses molécules pré biotiques se mettent assez facilement en boule, formant de petites sphères dotées d'une membrane. En effet, tout le monde sait que l'huile (un lipide) ne se mélange pas à l'eau. En agitant fortement les deux composés ont obtient une émulsion où l'huile forme de minuscules gouttelettes dans l'eau. Cela s'explique car certains lipides sont constitués de 2 parties :
- une tête hydrophile (qui aime l'eau, c'est à dire qu'elle possède une certaine affinité envers les molécules d'eau), constituée de glycérol,
- une queue hydrophobe (qui n'ont aucune affinité avec l'eau), constituée d'acides gras.
Cette association forme un phospholipide.
Les molécules d'eau par leurs propriétés physico-chimique tendent à se lier entre elles ou avec les molécules qui leur sont affines. La queue de phospholipides va ainsi être repoussée naturellement sous l'effet de la pression des molécules d'eau qui se réunissent. Avec l'agitation, cela entraîne le regroupement des phospholipides, et la formation de micelles, sorte de petites vésicules. A plus forte concentration se forme une émulsion constituée de globules assez gros. De la même les protéines peuvent aussi se protéger de l'eau par groupement en lamelles ou globules, notamment sous l'effet de hautes températures.
C'est à partir de ce phénomène chimique que les coacervats sont apparus. Ce ne sont que des sphères de lipides isolant une portion du milieu aqueux. Les phospholipides vont s'assembler sous forme de deux couches, une où les têtes des phospholipides font face au milieu extérieur, et une interne, où les têtes des phospholipides font face au milieu intérieur de la protocellule ou coacervat.
Cette "cellularisation" isole ainsi une petite portion de la soupe primitive, riche en molécules, qui va devenir le cytoplasme. Au début, ces coacervats étaient un désavantage car leurs molécules isolées du reste du milieu, les possibilités d'évolution étaient réduites. Mais à partir du moment où l'ARN est devenu capable d'engendrer des enzymes, dont certaines facilitaient sa propre réplication, ce désavantage s'est transformé en avantage.
Les propriétés des membranes lipidiques rappellent, en mieux, celles des bulles de savons. Ainsi sous l'effet d'une forte agitation les grosses cellules peuvent se diviser.
Rapidement les molécules hydrophobes, formées au hasard des mutations et des erreurs de réplication, ou de traduction, vont s'intégrer dans le seul milieu protégé de l'eau, la membrane lipidique. Certaines de ces molécules vont permettre un échange entre le milieu intérieur et extérieur, d'autres seront des enzymes fixées sur la face interne de la membrane cellulaire (c'est le cas notamment des enzymes permettant la formation des phospholipides).
Le CMP, qui participe entre autre à la formation de l'ARN, va permettre la formation de polysaccharides, de longues chaînes de sucres. Dans un premier temps ces molécules vont servir de réserves, puis certaines d'entre elles vont être intégrées à la membrane, lui conférant de nouvelles propriétés :
- Possibilités d'échanges des polysaccharides avec le milieu extérieur,
- Renforcement de la membrane, puis formation d'une paroi, grâce à l'apparition de muréine ou peptidoglycane un polymère de glucides et de peptides. La paroi forme une sorte de bouclier perméable pour la cellule : elle lui confère une grande rigidité sans trop entraver les échanges moléculaires. Cette paroi va limiter la taille des cellules et ainsi limiter leurs besoins en éléments extérieurs.
Les protéines membranaires vont peu à peu se spécialiser :
- certaines vont former des pores permettant divers échanges de petites molécules,
- d'autres vont jouer le rôle de transporteurs pour le passage d'une substance plus ou moins spécifique à travers la membrane (à la façon des portes tourniquets de certains magasins). Ce transport pourra être "gratuit", ou nécessiter l'échange d'une autre substance, qui prendra le chemin inverse, ou encore avoir besoin d'énergie (consommation d'ATP) pour fonctionner, on parle alors de transport actif.
Ces différents transports vont permettre de contrôler les entrées et sorties de la cellule, et ainsi de favoriser l'adaptation de la cellule vis à vis des modifications du milieu extérieur, en rejetant ou concentrant certains molécules.
L'apparition d'un transporteur de protons (H+) va être un avantage indéniable pour la cellule. Deux types de transporteur remplissent ce rôle. Un qui permet en recevant un électron d'une molécule extérieure, de rejeter un proton, et un autre qui rejette les protons en consommant de l'ATP. On appelle ces systèmes des pompes à protons. Quel que soit le transporteur le rejet de protons permettra de maintenir le milieu interne à un pH neutre (en y supprimant l'acidité due aux protons), tout en créant un gradient (ou potentiel) de protons à l'extérieur. Et si besoin est, le phénomène inverse est possible.
La molécule d'ARN a rapidement évolué vers un modèle stable dont l'évolution devient difficile. Un nouveau type de sélection s'est fait jour : certains ARN sont capables de se lier à un acide aminé. Une telle association les protège mieux de la dégradation en les rendant plus compact. Devenant de plus en plus nombreux, il leur est alors possible de se lier entre eux et de former de petits peptides par association de leur acide aminé. C'est là qu'interviennent des ribozymes, capables de catalyser ce type de liaison, et une troisième molécule d'ARN, qui par sa séquence de nucléotides dirige l'ordre de formation du peptide (elle fait alors office de gène). La fonction de traduction était née.
Là encore parmi les peptides formés certains possèdent une activité catalytique et préfigurent les futurs enzymes. Certains ont pu faciliter la réplication des ARN par exemple. C'est à ce stade que l'apparition de la cellule est devenue nécessaire. Si de telles enzymes étaient libres dans le milieu, elles profitaient à tous les ARN concurrents, ne permettant pas à l'ARN qui les codait de se mettre en valeur par rapport aux autres, ce qui aurait probablement entraîné tôt ou tard sa disparition. Si l'ARN et l'enzyme étaient isolés dans une protocellule, celle-ci serait grandement avantagée car elle seule bénéficiait des produits de la traduction.
Sous l'action d'enzymes, certains ARN se combinent entre eux, et forment ainsi un ARN plus long et donc des peptides plus longs. Cette "combinaison" ou épissage a pu être réalisé grâce à l'action de certains ribozymes. Parmi ces nouveaux peptides, certains apportent de nouvelles propriétés. Ainsi une enzyme permettant de fabriquer l'ADN a pu voir le jour. Cette transcriptase reverse comme on l'appelle actuellement permet à partir d'une molécule d'ARN (d'un seul brin) de créer une molécule double brin d'ADN. Les différences entre ARN et ADN (Thymine à la place de l'uracile, désoxyribose à la place du ribose) peuvent provenir de l'affinité de la transcriptase reverse pour de telles molécules.
Une telle enzyme devait en fait permettre à la fois la formation d'ADN à partir d'ARN, la réaction inverse et la réplication. Puis avec le temps de nouvelles formes plus spécialisées l'ont remplacé.
L'avènement de l'ADN a permit de stocker les gènes primitifs (ex ARN) en un seul exemplaire au lieu d'en nécessiter un grand nombre, elle a également favorisé leur stabilité.
L'hypothèse d'un monde de l'ARN, à l'origine de toutes les autres molécules du vivant (protéines, ADN) est séduisante. Mais il convient de rappeler que l'ARN est très sensible à la chaleur. On peut donc supposer que l'ARN c'est formé au niveau des sources hydrothermales mais que sa survie provient d'une migration vers des eaux moins chaudes.
Conjointement aux développements de toute cette chimie, les fameuses "conditions pré biotiques" qui régnaient sur Terre il y a 4 milliards d'années ont fait l'objet de nombreux travaux. Oparine pensait que la chimie de la vie avait débuté dans l'atmosphère, réservoir de petites molécules gazeuses contenant notamment du carbone, élément omniprésent dans les molécules du vivant. Il imaginait qu'elle se poursuivait dans les océans où l'eau liquide joue un rôle de solvant déterminant. C'est avec une atmosphère riche en méthane (CH4) que les réactions chimiques prébiotiques marchent le mieux. Mais dans ces conditions, la température aurait été trop basse et l'eau aurait gelé à la surface de la Terre. C'est pourquoi, une atmosphère riche en gaz carbonique (CO2) paraît incontournable, car elle aurait confiné assez de chaleur à la surface terrestre pour que l'eau, le premier solvant, reste liquide. Le problème, c'est qu'une atmosphère riche en gaz carbonique ne favorise pas la chimie pré biotique.
L'atmosphère est-elle le berceau de la vie ? Deux candidats se proposent actuellement pour assurer la relève : les sources chaudes sous-marines (si elles existaient alors) qui réunissent beaucoup de conditions propices à la chimie pré biotique, ou bien l'espace... !
De nombreux corps cosmiques, micrométéorites, grains de poussière, comètes... sont, en effet, riches en molécules fort intéressantes pour les réactions pré biotiques. Depuis toujours, ils ont largement disséminé leur matière sur la Terre. De quoi y ensemencer la vie. Mais aussi de nous faire rêver bien plus loin, d'imaginer que la vie elle-même nous vient de l'espace, et que nous ne sommes pas seuls dans l'Univers...
Les molécules organiques ne s'associent pas en n'importe quel endroit de leur structure, elles possèdent des groupements d'atomes plus réactifs que d'autres où l'association aura lieu. Donc les associations ne sont pas totalement dues au hasard mais sont "dirigées" en fonction de la présence ou non de tels sites réactifs.
Quand deux acides aminés se rencontrent au hasard, l'extrémité la plus "acide" (en rouge) d'un des acides aminés va s'associer à l'extrémité la plus "basique" de l'autre molécule (en bleu), avec une libération d'eau.
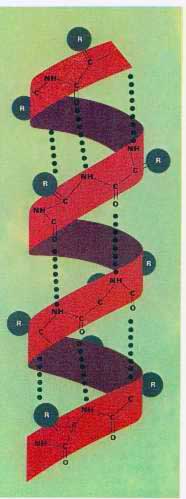
C'est ce que l'on appelle une liaison peptidique. Cette liaison est à l'origine de la formation de très grosses molécules, les protéines, par chaînage de différents acides aminés à la suite les uns des autres. Dans la soupe primitive, les protéines ne pouvaient pas être formées en raison de l'absence d'enzymes suffisamment élaborées, mais de petits peptides ont pu se former de cette façon sous l'influence de fortes températures.
Ces longues molécules se replient dans les trois dimensions, par regroupement d'acides aminés distants, en fonction d'affinités interne. La configuration 3D des protéines leur a apporté une stabilité plus grande, mais a également permit l'apparition de nouveaux sites réactifs par rapprochement de 2 séquences d'acides aminés qui étaient jusqu'alors séparées. Ainsi certaines protéines ont eu une activité catalytique sur d'autres molécules, ce sont les enzymes. Elles permettent de découper, coller, modifier les molécules sur lesquelles elles agissent.

La stabilité des molécules permet une meilleure résistance aux conditions environnantes. De même la forte synthèse d'une molécule permet de contrecarrer son taux de destruction. Donc les réactions de catalyse en chaîne ou d'autocatalyse permettent une meilleure stabilité.
Les éléments ne pouvant faire face aux divers agents destructeurs (pH, radiations, température, potentiel d'oxydoréduction etc...) seront détruits et leurs déchets serviront à la synthèse de nouvelles molécules, ainsi de suite, et seules les molécules stables resteront.
Quoi qu'il en soit, les briques de la vie ont pu être créées... Les briques, oui, mais la vie ??? Comment créer la vie ???
Le hasard ??? Non, ce serait impossible... autant vouloir construire une maison en remuant quelques millions de briques dans une immense cuve de béton liquide... impensable. Réfléchissons... "Si je veux une poule, il me faut un œuf, mais pour avoir un œuf, il me faut une poule... et je peux retourner le problème dans tous les sens dans mon esprit, la vie ne peut être issue que de la vie. La question que posait le regretté Raymond Devos est encore d'actualité : qui de l'œuf ou de la poule est apparu le premier ?" Personne ne le sait encore... mais des modèles biologiques existent essayant d'expliquer le comment du pourquoi. Voici où en est le modèle le plus usité actuellement.
Pour qu'il y ait une vie, il faut qu'il y ait des protéines et pour qu'il y ait des protéines, il faut des acides aminés. Les acides aminés sont des molécules essentielles à la vie, ce sont les briques de la vie. Elles sont formées d'atomes de carbone, d'hydrogène, d'azote et d'oxygène.
Des acides aminés, on en trouve partout : dans l'eau de mer, dans les lacs et les rivières et même sur les météorites et les comètes... c'est dire si ces atomes se lient facilement... d'ailleurs, l'expérience de Miller l'a prouvé. Dans de telles conditions, on peut dire que l'apparition de la vie était inévitable. Il a suffit d'attendre assez longtemps (dans le cas de la Terre, il a fallu moins d'un milliard d'années, ce qui est somme toute assez rapide !!) pour que des associations d'acides aminés, simplement générées par les lois physiques qui régissent les atomes et les molécules, donnent après des milliers, voire des millions d'essais infructueux, une petite chose réagissant sur le milieu : une petite sphère, un petit coacervat isolant une petite partie de la soupe primitive riche en matière organique : LA VIE.
Il parait logique que seules les protocellules capables de survivre ont eu un intérêt dans l'évolution. Ces coacervats étaient capables de se maintenir en vie en sachant puiser les éléments qui leur étaient nécessaires dans le milieu mais aussi en éliminant les déchets de leur métabolisme pouvant leur être nocif.
Au début les protocellules devaient tirer l'énergie nécessaire à leur "métabolisme" dans le milieu extérieur, en absorbant les molécules d'ATP ou de thioesters.
Mais ces molécules étaient difficilement régénérables. Les coacervats avaient également besoin de molécules pour les dégrader et récupérer soit de l'énergie, soit des éléments nécessaires à la bonne marche des réactions. Pour cela les transporteurs membranaires étaient importants, notamment les protéines canaux. Certains coacervats ont ensuite acquis la possibilité de rejeter les déchets, molécules nocives à la cellule, ainsi que des enzymes. Celles-ci en dégradant les grosses molécules du milieu extérieur offrent alors une nouvelle source de nourriture aux protocellules. Il va sans dire que les coacervats possédant les "gènes" permettant cette fonction de digestion extracellulaire ont rapidement supplanté les autres. Les cellules à paroi furent plus avantagées que les autres car leurs enzymes, bloquées entre la membrane et la paroi, restaient à disposition quelle que soit l'agitation du milieu tandis que les autres cellules devaient être en milieu stable car sinon enzymes et résultats des dégradations auraient été emportés par le courant.
La réaction de fermentation permet de consommer des molécules comme des glucides pour en retirer une certaine énergie (stockée sous forme d'ATP) qui sera utiliser pour d'autres réactions. Au début ces réactions devaient être simples et peu productives, puis peu à peu elles ont évolué vers la glycolyse actuelle.
Les protocellules qui ont renfermé les enzymes nécessaires à la fermentation ont pu s'affranchir en partie de la recherche d'énergie. Ce n'était toutefois pas suffisant pour une véritable autonomie, elles devaient trouver suffisamment de substrats pour satisfaire leurs besoins.
Les protocellules ayant acquis la possibilité de régénérer de l'ATP à partir du système des pompes à protons vont être considérablement avantagées. En effet si une cellule possède à la fois une pompe à protons fonctionnant avec des électrons, et une fonctionnant par transport actif, il est alors possible que les protons rejetés par la première, alimentent la seconde, qui au lieu de consommer de l'ATP va en régénérer.
Rapidement ce système s'est perfectionné, et plusieurs transporteurs d'électrons se sont regroupés au sein de la membrane. Chez certaines protocellules des pigments, capables d'absorber les rayonnements solaires (en particulier les UV), se sont associer à de telles chaînes de transporteurs d'électrons. Ainsi les électrons libérés par le pigment sous l'effet des rayonnements sont pris en charge par la chaîne de transporteur, ce qui permet une forte production d'ATP. La cellule a pu ainsi se libérer complètement de la source d'énergie chimique (issue de la fermentation) en la remplaçant par une source infinie (ou presque) : la lumière. La cellule est alors capable de vivre dans des milieux très pauvres en molécules organiques grâce à cette autotrophie. La colonisation de toutes les zones sans "vie" des océans est rapide.
Si une cellule possède des propriétés avantageuses, celles-ci ne seront vraiment utiles que si elles sont transmises à la descendance. Les propriétés de la membrane cellulaire font que la division des cellules est assez simple, et peut se faire sous l'action d'une simple agitation du milieu. Dans ce cas, le contenu de la cellule est distribué dans les deux nouvelles cellules de façon totalement aléatoire. Cette division est nécessaire car si la cellule croît sans limite, son volume devient trop important et ses capacités d'absorption trop faible. Une hétérogénéité dans la membrane peut également provoquer un bourgeonnement au cours de la croissance cellulaire. Actuellement on ne connaît pas le mécanisme de division qui a pu affecter les cellules entourées d'une paroi.
Avec l'apparition de l'ADN, les divisions de ce type devaient souvent engendrer une cellule complète et une sans matériel génétique, mort-née. Puis l'ADN s'est fixé un jour à la membrane ainsi que les enzymes nécessaires à sa réplication. La division a pu alors se faire de concert avec la réplication, permettant la formation de deux cellules à peu prés identiques, chacune possédant son ADN. Ce système a ensuite évolué vers le modèle du chromosome bactérien.
La transformation de complexes de protéines membranaires en cils ou flagelles mobiles ont permit aux cellules de se déplacer rapidement, et donc de trouver de nouveaux milieux plus riches, mais aussi de favoriser les rencontres entre cellules. La modification de cils en pili sexuels a permit les échanges d'ADN (conjugaison) et favorisé les recombinaisons.
La capacité à contrôler leur métabolisme est nécessaire aux cellules. Parmi la grande diversité des protéines transmembranaires synthétisées, certaines changent de conformation lorsqu'une molécule spécifique vient à leur rencontre. Ce changement de conformation peut permettre l'apparition d'une activité enzymatique de la protéine du coté cytoplasmique qui va modifier, directement, ou indirectement, une autre protéine transmembranaire. Ainsi la membrane voit l'apparition de récepteurs qui contrôlent l'ouverture ou la fermeture d'une protéine canal par exemple. La cellule peut alors contrôler ses échanges, en fonction de la richesse du milieu extérieur.
De même au sein du cytoplasme certains produits de réactions vont agir en activant ou inhibant d'autres réactions, ou même agir directement sur les gènes. Petit à petit, l'ensemble des réactions de la cellule sont en relation et s'autorégulent.
Ainsi parmi les cellules primitives certaines deviennent autotrophes tandis que les autres restes hétérotrophes et sont dépendantes des premières pour en récupérer les molécules organiques. Avant cette séparation, ou peu après, débute la longue histoire de l'évolution biologique et de la vie. Les protocellules ne peuvent plus être considérées comme de simple réservoir à réactions biochimiques mais comme de véritables unités de la vie, les cellules (qui ont les facultés d'autoreproduction d'autoconservation et d'autorégulation).
La vie était donc inévitable, inéluctable, car des lois physiques et chimiques qui régissent les atomes au sein de la soupe primitive ont eu tout le temps d'essayer, à tâtons, par essais et erreurs, je le conçois, toutes les possibilités de liaisons entre atomes et ainsi d'arriver à un être vivant.
Pour me contacter, me faire part de vos idées, me poser vos questions, me laisser vos remarques...
cliquez ici :

![]()
Retour vers le sommaire
![]()